Jakie informacje z bodźców do nas docierających są istotne?
Jakie możemy wykorzystać? Co było biologicznie możliwe a co nie?
Dlaczego mózg zbudowany jest tak a nie inaczej?
Dlaczego mamy wzgórze i co robi wstępne przetwarzanie informacji zanim dotrze ona do kory?
Widmo elektromagnetyczne: fizyka determinuje, co da się odbierać a co nie. Za długie fale nie wywołują dostatecznie silnej reakcji cząsteczek chemicznych i tkanek, za krótkie wywołują raka. Jak biomolekuły reagują na promieniowanie i jak ta reakcja może się rozprzestrzenić w materii biologicznej?
Istnieje kompromis pomiędzy stopniem naprawiania komórek i ich możliwością zmienności.
Analizując percepcję musimy się zastanowić, czy nasza fenomenologia pasuje do fizycznych podstaw powstawania wrażeń, a więc rozchodzenia się aktywacji neuronalnej w mózgach.
Na co reagują pola recepcyjne kory zajmującej się analizą danych zmysłowych? Nawet najprostsze bodźce - z punktu widzenia naszych wyobrażę - wywołują złożone reakcje mózgu. Co jest dla mózgu proste? Czy są takie bodźce, które wywołują powtarzalną, zlokalizowaną reakcję?
Istnieje kompromis pomiędzy liczbą słów danego języka, możliwościami kompozycji tych słów pozwalającymi na opis obiektów i zdarzeń, oraz dokładnością tego opisu.
Emocje w każdym języku opisywane są w nieco inny sposób, chociaż cała przestrzeń opisu to te same reakcje afektywne, różniące się nieco w zależności od organizmu i kultury, ale w sumie podobne. W przypadku percepcji opis wydaje się bardziej jednoznaczny, chociaż za tymi samymi słowami też stoją zróżnicowane wrażenia.
Kolor, wysokość i barwa dźwięku, zapach czy smak są perceptami, a nie fizycznymi własnościami fal elektromagnetycznych, akustycznych lub pobudzeń chemicznych docierających do zmysłów.
Mózg wydobywa z pobudzeń kory analizującej dane zmysłowe niezmiennicze własności, widzimy te same obiekty niezależnie od kąta, odległości i obrotu, lub kolory niezależnie od oświetlenia.
International Society for
Group Theory in Cognitive Science zajmuje się zastosowaniem teorii group, poszukiwaniem symetrii w procesach związanych z percepcją, głównie percepcją wzrokową.
Czy można opisywać świat za pomocą całkiem odmiennej siatki pojęciowej, podobnie jak to się dzieje z emocjami?
Jak to by wpłynęło na nasz sposób przeżywania wrażeń?
Istnieje kompromis pomiędzy liczbą słów danego języka, możliwościami kompozycji tych słów pozwalającymi na opis obiektów i zdarzeń, oraz dokładnością tego opisu i funkcjami komunikacyjnymi.
Używane pojęcia są zapewne wynikiem optymalizacji złożoności całego systemu: liczby zapamiętanych nazw do których możemy porównać nowe obiekty czy zdarzenia - np. cytryna jest jak mandarynka, tylko bardziej wydłużona i żółta - oraz prostych cech, które możemy użyć do opisu.
Badania fonologii języków pokazały, że pomimo różnej szybkości mówienia (japoński i hiszpański ma najwięcej sylab na sekundę, mandaryński i niemiecki najmniej) ilość informacji zawarta w wypowiedziach na jednostkę czasu jest podobna (Pellegrino i inn 2011), pomimo zmian powierzchniowych cały system wykazuje podobny stopień optymalności.
Patrząc na mandarynkę opisuję swoje wrażenia używając zarówno prostych perceptów - pomarańczowa, okrągła - jak i złożonych perceptów - pomarszczona i wyschnięta skóra, widać na niej zapadnięte bruzdy.
Neurofenomenologia powinna lepiej powiązać fizyczną aktywność obszarów w mózgu z subiektywnie postrzeganymi wrażeniami.
Używając podstawowych pojęć dotyczących percepcji mamy trudności ze zrozumieniem, co właściwie robi wtórna kora zmysłowa, na jakiego rodzaju bodźce reaguje - byłoby ciekawe zacząć od poszukiwania przykładów bodźców wywołujących podobne reakcje tych obszarów i zrozumienia, jakiego rodzaju złożone pojęcia są przydatne do ich opisu.
Odczuwanie bodźców można podzielić na cztery kategorie:
Czucie głębokie i trzewne jest często mało precyzyjne (protopatyczne); integracja infromacji odbywa się na głównie w pniu mózgu i ośrodkach podkorowych.
Czucie powierzchniowe i teleceptywne pozwala na precyzyjne (epikrytyczne) różnicowanie bodźców, precyzyjna analiza informacji odbywa się dodatkowo w korze mózgu.
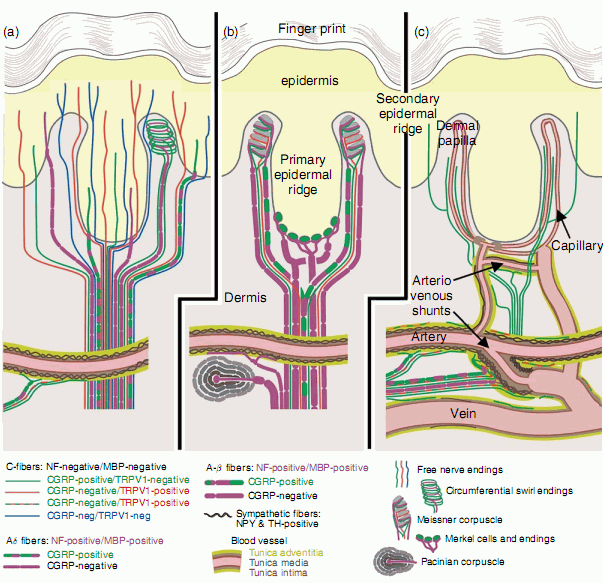
Mamy około 2 mld receptorów czuciowych. Jaka jest ich czułość?
Dzięki liniom papilarnym możemy odczuć nierówności rzędu 50 nanometrów,
czyli 5/100 000 mm!
Możemy też zauważyć różnice temperatury rzędu 0.01 stopnia Celsjusza.
Czucie powierzchniowe i kontrola ruchów wykorzystuje podobne mechanizmy: mapy somatosensoryczne i mapy ruchowe.
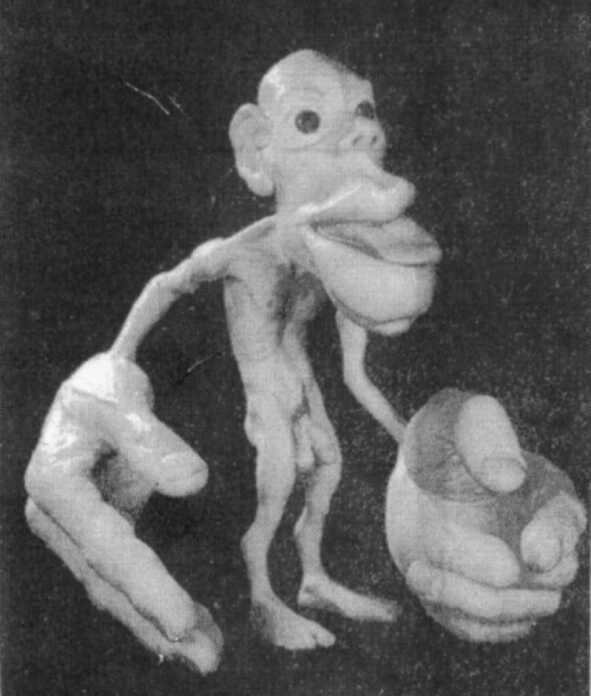
Wielkość obszaru analizującego dane zmysłowe zależy od ich przydatności dla danego zwierzęcia.
Gryzonie mają dużą powierzchnię poświęconą wibrysom.
U człowieka dominuje reprezentacja rąk i języka.

Informacja somatosensoryczna (czucie ciała) - obszar SI z tyłu bruzdy centralnej.
Informacja przekazywana jest od receptorów: dotyku, bólu, temperatury, wibracji, położenia kończyn, przez nerwy czuciowe do wzgórza i kory SI.
Szlaki czuciowe (film, tylko lokalnie).
Różne pobudzenia docierają różnymi nerwami: nerwy przekazujące sygnały szybko sygnalizują ból ostry, a wolno ból piekący.
Blokowanie wzajemne przepływu różnych informacji może nastąpić w rdzeniu, dlatego ucisk pomaga zmniejszyć wrażenie bólu.
Korelacje ruchu i czucia: pobudzenia z przeciwległych obszarów SI dochodzą do pobliskich obszarów MI (za bruzdą centralną).
Twarz i język reprezentowane są częściowo po tych samych stronach, a pozostałe części ciała przeciwlegle.
Zniszczenie kory SI powoduje zanik wrażeń czuciowych, jednak ból i temperatura po pewnym czasie pojawiają się; zgrubne rozróżnianie tych wrażeń możliwe jest już na poziomie wzgórza.
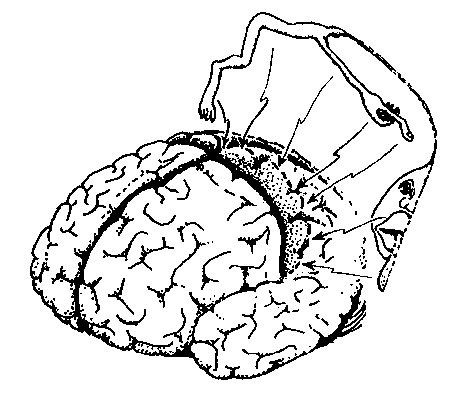
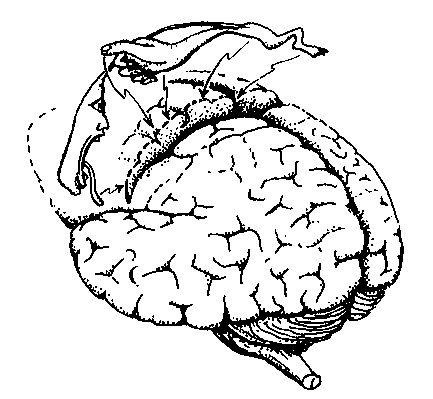
Różne części wzgórza zaangażowane są w projekcje somatosensoryczne:
VPL=Ventral Posterior Lateral Nucleus = jądro brzuszno-tylne boczne.
VPM = Ventral Posterior Medial Nucleus = jądro brzuszno-tylne przyśrodkowe.

|
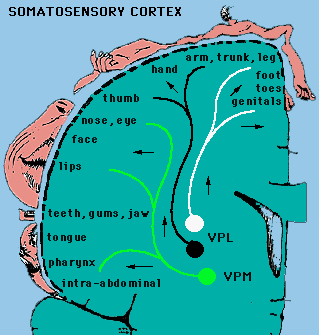
Mapy czuciowe: pobudzanie kory czuciowej słabym prądem wywołuje wrażenia dotyku, łaskotania, swędzenia.
Pobudzanie kory z przodu bruzdy centralnej wywołuje zachowania ruchowe (całe wyuczone ruchy). Drobne różnice pomiędzy mapą ruchową i czuciową. Obszary ciała mają podobne odwzorowanie w zakrętach kory, ale poza tym mamy duże indywidualne różnice i plastyczność (zmienność w czasie). Kora SI ma budowę kolumnową (baryłkową): na pobudzenia reaguje cała kolumna. Kora SI ma 4 podobszary Brodmanna, obszar 2 i 3a analizuje sygnały proprioceptywne, obszar 3b dotykowe, obszar 1 mieszane. Proprioceptywne - reakcje na ruch w określonym kierunku. |
 |
Obszar kory poświęcony analizie jest proporcjonalny do wagi bodźców. Celem analizy jest precyzyjna dyskryminacja.

Aplet pokazujący połączenia z korą czuciową.
Plastyczność map czuciowych: do lat 1970 sądzono, że mózg nie zmienia się po osiągnięciu dojrzałości.
Paul Bach-y-Rita
prowadził pionierskie prace już w latach 1960, pokazując jak można doprowadzić do rehabilitacji mózgu po udarze.
Doświadczenia z małpami
Michaela Merzenicha oraz
Edwarda Tauba .
Zmiana wielkości obszaru kory na skutek stymulacji lub braku bodźców (po uszkodzeniu nerwu lub amputacji palca).
Związanie kilku palców tak, że moga robić ruchy tylko jednocześnie, powoduje powstanie jednej rozległej mapy.
Reorganizacja map czuciowych i ruchowych po uszkodzeniu i zrośnięciu nerwów w odmienny sposób.
Symulacje rozwoju map topograficznych po urazach mogą być przydatne w rehabilitacji: metoda wymuszania ruchu przez ograniczanie (Constraint-induced movement therapy) Tauba przynosi bardzo dobre rezultaty.
Historia rozwoju ideii
neuroplastyczności
opisana jest w znakomitej książce:
The brain that changes itself.
Kryminalna historia Silver Spring Monkeys warta jest zastanowienia.

|
Przykład plastyczności: uczenie się rozpoznawania palców u nóg.
Czy naprawdę wiemy, co czujemy? Działanie neuronów nie zawsze jest precyzyjne: który palec u nogi został dotknięty? Jeśli go nie widzimy dokładność odpowiedzi osiąga 80-90%. Korelacja wzrok-dotyk szybko podnosi dokładność do 98%. Chodzenie w butach po paru dniach obniży dokładność do początkowej. Bez dyskryminacji nie potrafimy określić swoich wrażeń - jest to podstawa świadomej percepcji. |
 |
Dylemat plastyczności-stabilności jeszcze raz.
Za duża stabilność => brak adaptacji; za duża plastyczność => katastroficzne zapominanie, koniecnzy jest kompromis na każdym poziomie!
Neurony: stabilność synaps i zmiany umożliwiające uczenie się.
Mózg: uczenie się nowych faktów czy zachowanie stabilnego, usatlonego obraz świata.
Społeczeństwa: stabilność (partie konserwatywne) czy zmiany (partie postępowe)?
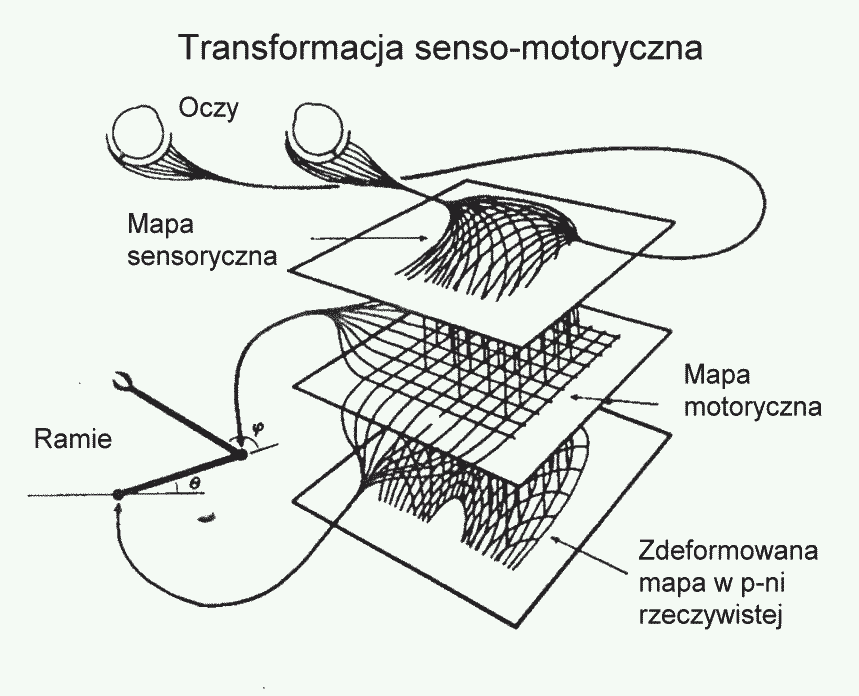
Po co nam w mózgu mapy topograficzne?
Umożliwiają szczegółową analizę sygnałów zmysłowych.
Transformacje senso-motoryczne wymagają złożonych, nieliniowych przekształceń, mapy to umożliwają.
|
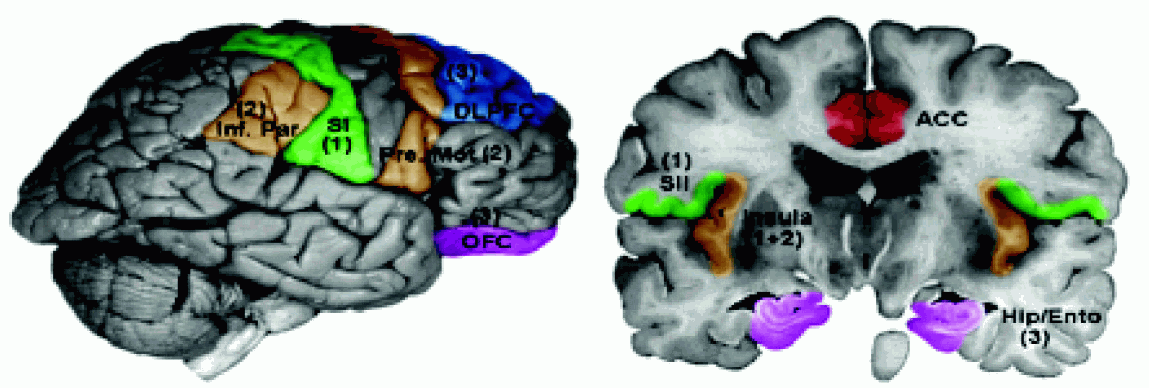
Wtórna kora czuciowa SII leży w głębi bruzdy środkowej i bruzdy bocznej (BA 43), w pobliżu kory wyspy, chociaż niektórzy zaliczają do SII również płacik ciemieniowy dolny (obszar BA5), nieco z tyłu w stosunku do SI.
Obszar ten wyróżniono tylko u ludzi i małp naczelnych, jest to obszar kojarzeniowy dla wrażeń dotyku i bólu. Komórki pobudzane są przez sygnały z większych obszarów ciała, z obu stron, bezpośrednio ze wzgórza, obszarów kory SI oraz spoidła wielkiego. Integracja informacji z obu stron ciała oraz hipokampu pozwala na uczenie się rozpoznawania przez dotyk i transfer tej umiejętności z jednej ręki do drugiej. |  |
|
Uszkodzenia SII i pobliskiej kory wyspy powodują
asymbolię bólu,
brak cierpienia (reakcji psychicznych) na ból, lub wywołują
stały ból piekący (kauzalgia).
|  |
|
Środki przeciwbólowe obniżają aktywność kory przedniego zakrętu obręczy.
|
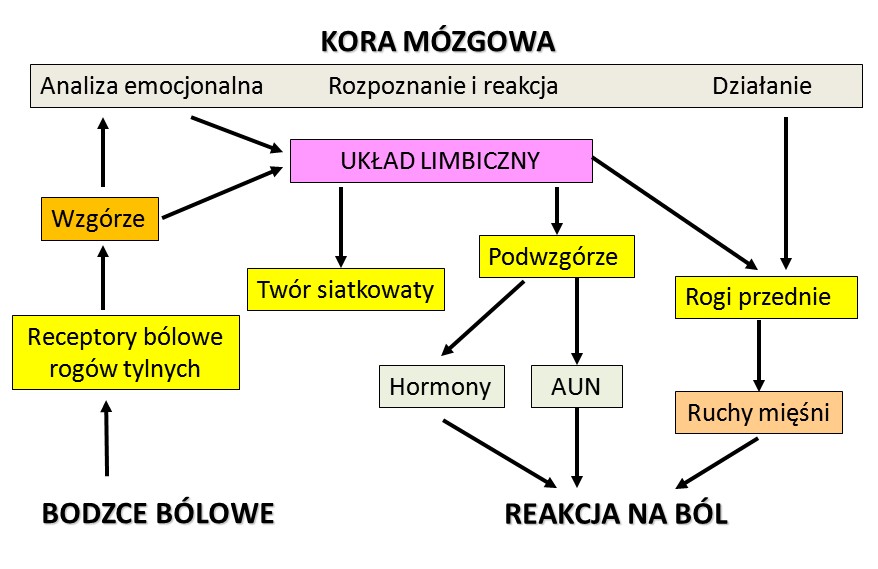
Uproszczony schemat dróg reakcji na ból.
|
SSA, dodatkowa kora czuciowa, znajduje się w płaciku ciemieniowym górnym (obszary 5 i 7 Brodmanna).
|
 |
Kora czuciowa ma trochę neuronów ruchowych (tylko dwie synapsy dzielą ją od kontroli mięśni), a kora ruchowa neuronów czuciowych.
Czucie zewnętrzne to analiza sygnałów z narzadów zmysłów.
Czucie głębokie to analiza sygnałów proprioceptywnych z mięśni, stawów, ścięgień, błędnika, informujące o położeniu i stanie ciała.
Czucie wewnętrzne, interoceptywne, analizuje informację z gruczołów i trzewii.
Agnozja (gr. a=negatywne, gnosis=wiedza) to niezdolność lub zaburzenie rozpoznania lub identyfikacji informacji zmysłowej.
Astereognozja to niezdolność do rozpoznania przedmiotów za pomoca dotyku.
Jest niezależna od zaburzeń czucia, upośledzenie rozpoznawania kształtu przy prawidłowym rozpoznawaniu wielkości, faktury.
Większość agnozji należy rozpatrywać na wyższym poziomie analizy powyżej kory analizującej dane zmysłowe.
Optyczna ataksja to niezdolność do chwytania przedmiotów pomimo prawidłowej analizy wzrokowej, np. nie można wlać wody do szklanki.
Uszkodzenia obszarów leżących w głębi przyśrodkowej kory ciemieniowej (LIP) wywołuje niezdolność do chwytania przedmiotów, np. lecącej piłeczki.
Obszar LIP zawiera topograficzną mapę obiektów istotnych z punktu widzenia bieżących możliwości działania, odwołania do specyficznych ruchów ciała i oczu, kategoryzację obiektów z punktu widzenia możliwych działań, spodziewane nagrody. LIP odpowiada więc za integrację informacji wzrokowo-przestrzennych, poznawczych i ruchowych, z mechanizmami uwagi.
Połączenie kory SI i SSA umożliwiają przepływ informacji pomiędzy lokalizacją wzrokową a pobudzeniem czuciowym.
Przykłady:
Utrata propriocepcji w wyniku zapalenia nerwów prowadzi do ślepoty ciała!
Opis przypadku:
"Bezcielesna kobieta", Oliver Sacks (w książce "Mężczyzna, który pomylił swoją żonę z kapeluszem").
Wystarczy informacja wzrokowa by kontrolować ciało, dzięki połączeniu obszaru PPC 5 i SI.
Kończyny fantomatyczneto silne odczucie obecności amputowanych kończyn, pojawiające się u 60-80% osób po amputacji.
Często jest to odczucie bolesne, czasami mogą sie pojawić wszelkie inne wrażenia czuciowe: ciepło/zimno, swędzenie, ucisk, łaskotanie, wrażenie ruchu i próba chwytania nieistniejącą kończyną.
Odpowiedzialna jest za to reorganizacja kory czuciowej wynikająca z pobudzeń z okolic ciała, które mają zbliżone pola projekcyjne w oszarze SI, słabe połaczenia normalnie maskowane przez odczucia płynące z brakującej części ciała.
Takie odczucia nie pojawiają się w przypadku porażenia kończyn dolnych (paraplegii).
Bóle fantomowe pojawić się mogą u osób po amputacji, z uszkodzonym rdzeniem, z wrodzonym paraliżem.

Ramachandran: chorzy z kończynami fantomatycznymi cierpią w nich silne bóle, np. odczuwają wrastanie paznokci w zaciśniętą dłoń.
Ból wywołuje stymulacja obszarów, których reprezentacje przylegają do kory odpowiedzialnej za wrażenia z brakującej części ciała; widać wtórna kora SII dostaje mylne sygnały.
Terapia jest możliwa przez pokazywanie im ruchu własnych zdrowych kończyn,
odbitych w lustrze lub obserwowanie ruchu czyjejś ręki.
Informacja wzrokowa pobudza tu korę SI w obszarze odpowiadającym utraconej kończynie i wywołuje reorganizację tej kory, bez takiej informacji nie dochodzi do reorganizacji.
Eksperyment z butem:
Dużo rzadszy przypadek: nadliczbowa kończyna fantomowa, której co prawda nie można użyć do manipulacji fizycznymi przedmiotami, ale można się nią podrapać! (Khateb i inn. 2009)
Kończyny niechciane (dysmorfia ciała): silny wstręt do własnego ciała, poczucie obcości, dość częste, 1:50 osób.
Skrajne przypadki zaczynają się w dzieciństwie, "życie w ciele, które nie jest własne".
Traktowana jako obsesja, pacjenci walczą o prawo
do amputacji kończyn (apotemnofilia).
Kilku osobom w Anglii obcięto kończyny (BBC Horizon
'Complete Obsession').
Co może być przyczyną?
Sprzężenie obszarów interpretujących informację z kończyn w obszarze SII z ciałem migdałowatym może wywołać nieprzyjemne odczucia.
Nie ma dobrych metod leczenia, można próbować pobudzania nerwów czuciowych w połaczeniu z przyjemnymi bodźcami.
Inne obszary zorganizowane topograficznie, w których pojawia się informacja somatosensoryczna:

Wzrok u zwierząt różnego gatunku realizowany jest na wiele sposobów: ślimak ma komórki światłoczułe bez soczewek,
owady złożone oko i 10-30.000 heksagonalnych fasetek,
ssaki mają
oko z siatkówką i soczewką,
głowonogi mają oczy podobne do ssaków.
Oko jest wysuniętą na zewnątrz częścią mózgu, "zwierciadłem duszy".

Oczy wykorzystują światło, powinny więc wyodrębniać ważne informacje; potrzebne są różne kompromisy:
Siatkówka ma złożoną budowę, niezbyt doskonałą:
Webvision, czyli wszystko na temat budowy oka, fotoreceptorów, siatkówki i trochę o korze wzrokowej.
Dane liczbowe na temat
siatkówki i jej połączeń.
Oczy głowonogów, np. ośmiornic są lepiej skonstruowane niż kręgowców: u człowieka światło dociera do czopków i pręcików przez trzy warstwy komórek i jest osłabiane.
Światłoczułe pręciki i czopki ośmiornice mają w przedniej warstwie, nie ma plamki ślepej. Inaczej działa też mechanizm skupiania, nie ma więc krótko i dalekowzroczności.
Komórki zwojowe wysyłają informacje o kilkunastu różnych "ścieżkach wideo" z których tworzy się w mózgu obraz świata.
Nawet dla prostego pobudzenia małego fragmentu siatkówki impulsem świetlnym 12 komórek zwojowych wysyła odmienne "ścieżki filmowe" wgłąb mózgu.
Obrazy na filmie pochodzą z neuromorficznego obwodu scalonego, który przetwarza informację w podobny sposób do siatkówki.
Każda grupa komórek filtruje specyficzne cechy obrazu: kontury, cienie, tekstury, oświetlone powierzchnie.
Każdy strumień informacji jest aktywny tylko przez milisekundy.
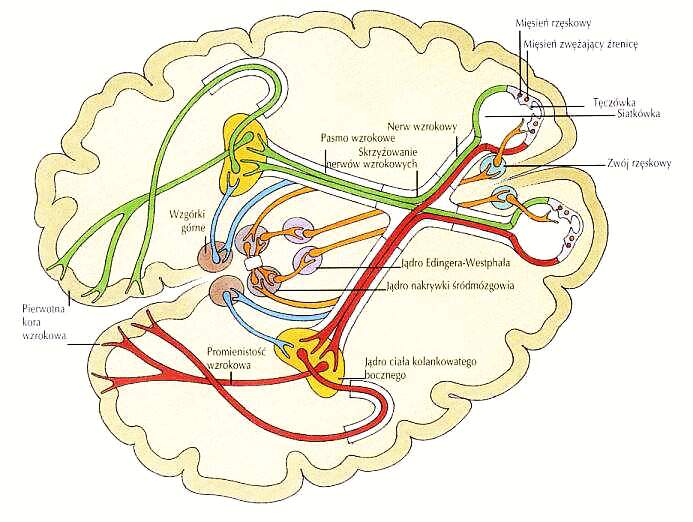
Szlaki wzrokowe: siatkówka, 6 mln czopków, 120 mln pręcików => ok. milion aksonów => ciało kolankowate boczne wzgórza => promienistość wzrokowa => obszar pierwotnej kory wzrokowej V1 => wyższe piętra układu wzrokowego => obszary kojarzeniowe i wielomodalne.
Szlak wzrokowy (film, tylko lokalnie).
Kora
obszaru V1, zwana jest również korą prążkowaną (białe paski na szarym tle, aksony promienistości wzrokowej kończące się w warstwie 4).
V1 zawiera komórki zorganizowane w kolumny dominacji dwuocznej i kolumny orientacyjne,
retinotopicznie (bliskie komórki reagują na bliskie sobie punkty w polu widzenia).
Proste komórki warstwy 4 reagują na paski o określonym nachyleniu, kontrastowe krawędzie, pobudzenia z jednego oka.
Znaczna część środkowego obszaru V1 reaguje na sygnały dochodzące od
okolic plamki żółtej (dołka środkowego) oka, gdzie gęstość receptorów jest największa.
Komórki złożone w pozostałych warstwach V2-V5 reagują na sygnały z obu oczu.
Prawidłowy rozwój
układu wzrokowego wymaga odpowiedniej stymulacji w dzieciństwie.
| Czemu nie widzimy obrazu odwrotnie?
Pryzmatyczne okulary odwracają obraz: kompensacja dla obrotu o 180 stopni zajmuje kilka dni. Sama forma tego pytania pokazuje, jak bardzo wierzymy w to, co widzimy, w istnienie homunculusa, który postrzega. W mózgu wszystko staje się elektrycznymi impulsami, nie istnieje góra czy dół. Reprezentacja świata jest tylko reprezentacją relacji, informacją potrzebną do działania w świecie. | 
|
Płat potyliczny: poniżej bruzdy ciemieniowo-potylicznej, wyraźnie widocznej na powierzchni przyśrodkowej.
Obszary Brodmana: 17 czyli V1, kora prążkowa, pierwotna kora wzrokowa.
Obszar 18, czyli V2, drugorzędowa okolica wzrokowa, połączona obustronnie z V1 i wysyłająca sygnały do wyższych pięter V3-V5.
Obszar 19, czyli okolice V3-V5 (strumień grzbietowy, w stronę kory ciemieniowej).
Im wyżej w hierarchii tym silniejsza jest modulacja aktywności związana z uwagą, w V1 jest ona słaba, w V4 silna.
Dwa strumienie informacji wzrokowej, Ungerleider i Mishkin (1982): istnieją dwa w znacznej mierze rozdzielone szlaki przetwarzania informacji wzrokowej, biegnącej już od oka.
Wielkoziarniste komórki PA siatkówki, 3 typy stożków fotorecepcyjnych, duże pola recepcyjne, szybko przewodzące aksony, pobudzenie dla światła w szerokim paśmie.
Drobnoziarniste komórki PB, 1 lub 2 typy stożków fotorecepcyjnych, małe pola recepcyjne, wolno przewodzące aksony, rozpoznają opozycje barw.

Szlak wielkokomórkowy: biegnie do dwóch wielkokomórkowych
warstw LGN (jest w nich ok. 100.000 komórek), charakteryzuje go niska rozdzielczość przestrzenna, wysoka wrażliwość na kontrast, szybkie przesyłanie sygnałów, bez informacji o kolorze.
Ta informacja trafia przez płat potyliczny
szlakiem grzebietowym do kory ciemieniowej.
Dochodzi do warstwy 4B w V1, stąd do grubych ciemnych pasków obszaru V2, analizuje informację o ruchu obiektu.
W V1, warstwa 4B => V5, lokalizacja w polu widzenia, ruch.
V5 pobudza płat ciemieniowy, PPC (tylna kora ciemieniowa), obszar 7 i 5; umożliwia to orientację przestrzenną, postrzeganie głębi i ruchu, połączenie z wzgórkami czworaczymi (orientacja oczu).
Szlak drobnokomórkowy ma 4 drobnoziarniste warstwy i 10 razy więcej komórek niż wielkokomórkowy w LGN.
Duża rozdzielczość przestrzenna, kolor, wolniejszy przesył informacji, niska wrażliwość na kontrast.
Ta informacja trafia szlakiem brzusznym do kory dolnoskroniowej.
V1 => V2 obszar międzyplamkowy, reaguje na orientację linii, daje dużą ostrość widzenia, bez koloru.
V1 => V3 obszar plamkowy, reaguje na kształty, reakcja na kolor w neuronach w ciemnych prążkach V3.
V2 => V4, główny obszar analizy koloru, informacja dochodzi do kory dolnoskroniowej (IT).
Obszar IT w płacie dolnoskroniowym ma neurony reagujące na złożone obiekty.
Demo: laboratorium Keiji Tanaka, RIKEN.
LGN ma tylko 10-15% pobudzeń z siatkówki, pozostałe 95-90% z kory wzrokowej.
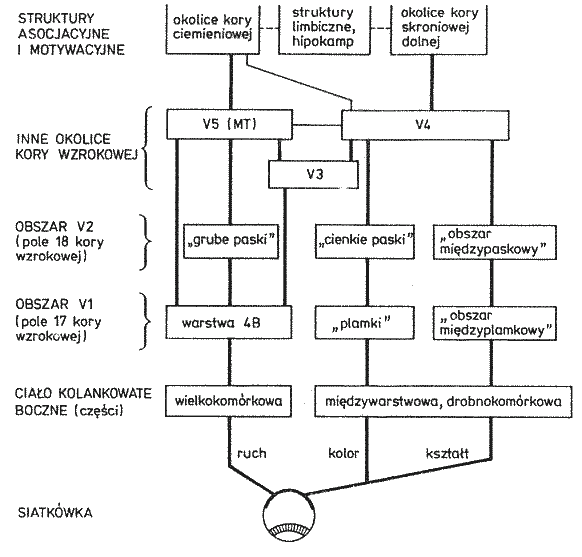
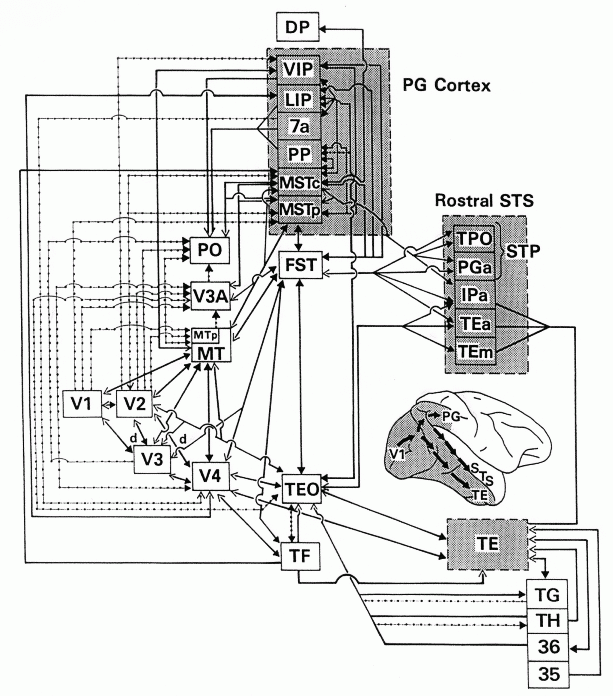
Szkic podobszarów układu wzrokowego.
Szkic dokładniejszy - makak
i jego rozpłaszczona kora.
Szkic dokładniejszy - człowiek
Mamy ponad 25 obszarów kory związanych z przetwarzeniem informacji wzrokowej, zorganizowanych w hierarchiczny sposób.
Agnozje wzrokowe.
Kiedy wszystko działa normalnie nie zauważamy, że stoi za tym złożona maszyneria, ale kiedy się coś popsuje ...
| Uszkodzenia (udary, wypadki, zmiany neurodegenercyjne) obszaru V2 wywołują zaburzoną percepcję kształtów. Agnozja kształtu ma wiele form: | 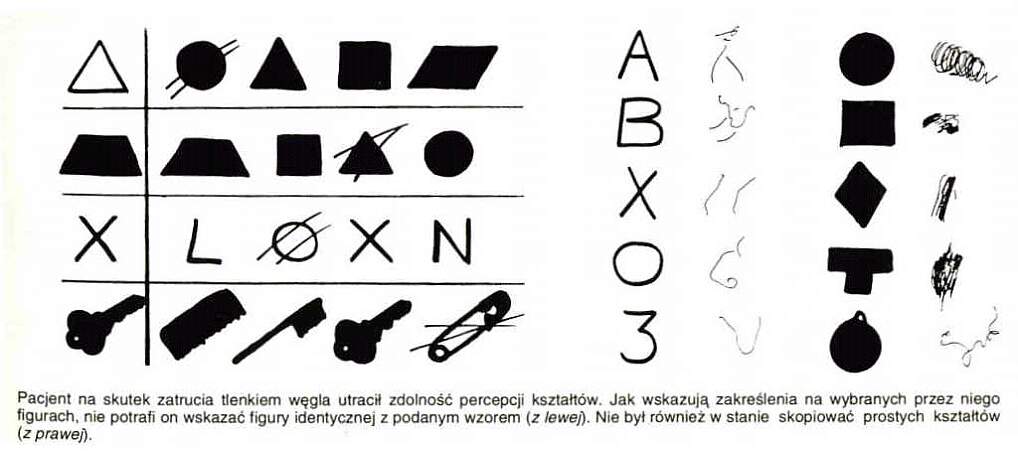 |
| Agnozja kształtu może być wybiórcza, np. bez prozopagnozji.
Pacjent widzi twarz ale nie poznaje warzyw, owoców i kwiatów, z których się ona składa. |  |
| Uszkodzenia obszaru V4 prowadzą do
mózgowej achromatopsji, ślepoty barw, czyli zaniku zdolności do widzenia kolorów.
Wrodzona achromatopsia jest wynikiem mutacji genetycznej prowadzącej do defektu czopków w siatkówce, może być endemiczna, np. na jednej z wysp Norwegii i Mikronezji (Sacks, The island of colorblind) większość społeczeństwa niezdolna była do widzenia kolorów. Anomia barw nie jest związana z percepcją ale zaburzeniami nazywania barw, są to uszkodzenia w obszarach trzeciorzędowych (zakręt kątowy). Anomia wzrokowa może przejawiać się trudnościami z uporządkowaniem kolorów, pomimo widzenia barw - brak zrozumienia koncepcji barwy? Anomia nazywania: brak skojarzenia nazwy z kolorem. | 
|
Uszkodzenia V5 -
akinetopsja, widać statyczne migawki, ale nie ruch, wrażenia przypominają widoki w świetle stroboskopwym.

Stymulacja zakrętu IT wywołuje halucynacje wzrokowe.
Obszar Brodmana 37, zakręt potyliczno-ciemieniowy (obok IT), rozpoznawanie twarzy i miejsc.
Uszkodzenia powodują
prozopagnozję, niezdolność do rozpoznawania twarzy; w tym obszarze ponad 90% komórek reaguje tylko na twarze.
Wszystkie twarze wydają się wówczas podobne (jak nam np. twarze chińczyków), ale można nawet nie odróżniać własnej twarzy od innych.
Czasami pomimo braku rozpoznania da się zaobserwować reakcje emocjonalne na poziomie podkorowym, np. reakcje GSR (czyli zmianę oporności skóry na skutek pocenia w wyniku takich reakcji).
Zachowana jest zdolność do rozpoznawania zwierząt (np. indywidualnych owiec w przypadku pasterza).
Normalnie układ wzrokowy się uczy, mózg przystosowuje się do nowych bodźców; w prozopagnozji nie potrafi się już nauczyć gdyż kora w tym się specjalizująca uległa uszkodzeniu.
 |
 |
| Analiza fMRI procesu rozpoznawania twarzy. | Widać wyraźną lokalizację aktywności w prawej półkuli, w zakręcie dolno-skroniowym (IT). Dokładne badania pokazały, że ponad 90% komórek w tym obszarze reaguje wyłącznie na twarze. Rozpoznawanie twarzy jest b. ważne z ewolucyjnego punktu widzenia. |
|
Uszkodzenia szlaku do IT i zakrętu kątowego prowadzą do
agnozji wzrokowej, czyli niezdolności do nadania sensu temu co się widzi.
Niemożliwe jest świadome rozpoznania przedmiotów przy zachowanej zdolności do działania, np. uchwycenia przedmiotu czy manipulacji nim (ślepota psychiczna). Zaburzenie dotyczyć może zdolności rozpoznawania obiektów wewnątrz jakiejś kategorii, np. samochodów, krzeseł, zwierząt czy palców ręki. Pomimo prawidłowego wykonania kopii rysunków pacjent nie ma pojęcia, co przedstawiają narysowane przedmioty. |

|
|
Uszkodzenia przepływu informacji wywołują liczne syndromy neuropsychologiczne.
Hipoteza Ungerleidera-Mishikna:
|

|
Sygnały wzrokowe przez wzgórki czworacze górne i wzgórek wzrokowy sterują sakadycznymi ruchami oczu.
Wzgórek wzrokowy ma połączenie z korą ciemieniową, przechowująca mapę umożliwiającą orientację w przestrzeni.
Obszar IT ma bezpośrednie projekcje z siatkówki, pozwalając na szybkie niedokładne pobudzenie najwyższych pięter układu wzrokowego: generuje to hipotezę wstępną - co widzimy?
Strumienie informacji zstępujące do niższych pięter pomagają w precyzyjnym rozpoznaniu: dopiero stany rezonansowe powstałe w wyniku pętli IT-V1 są uświadamiane.
Szybkie reakcje ruchowe jeszcze przed rozpoznaniem obiektu umożliwia szlak grzbietowy (kora ciemieniowa, a potem ruchowa).
Zamrożenie obszaru V5 u makaka powoduje niezdolność obszaru V2 do właściwej reakcji na docierające bodźce; nie ma hierarchii aktywacji w układzie wzrokowym, tylko współpraca każdego obszaru z innymi, sprzężenia są silne.
Świadomość wzrokowa, percepcja, zależy od pobudzenia szlaku skroniowego.
Milner i Goodale (1995): szlaki wzrokowe nie tyle określają co i gdzie, co umożliwiają działanie i percepcję.
Jest to uproszczenie, bo jest jeszcze stary szlak limbiczny, umożliwiający szybkie działanie w niebezpiecznych sytuacjach (po którym następuje fala strachu).
Potknięcie, szybkie odzyskanie równowagi i fala strachu to reakcja starego szlaku.
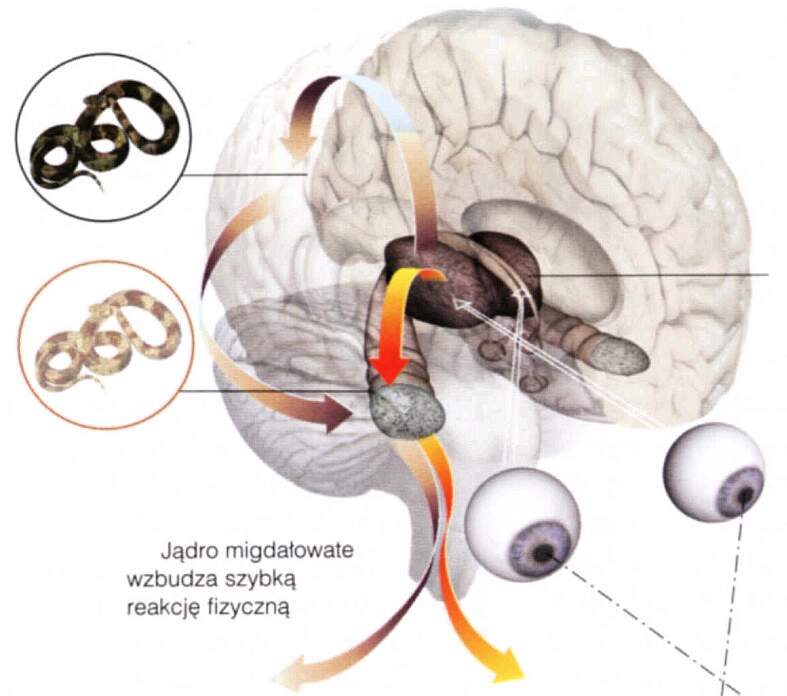
Wrażenia wzrokowe to funkcja wyższych pięter układu wzrokowego.
Pobudzenia z nerwu wzrokowego odpowiedzialne są za niewielką część aktywności powyżej V1 (ok. 10%).
We śnie możemy mieć wyraźne wrażenia wzrokowe bez pobudzenia siatkówki.
W jaki sposób tworzy się spójne wrażenie z aktywności różnych obszarów, obrazu rozbitego na różne elementy (kształt, kolor, ruch)?
Powstaje problem spójności wrażeń wzrokowych (visual binding).
Czym różnią się od siebie wrażenia z różnych zmysłów?
Symultagnozja (agnozja symultatywna): postrzeganie pojedynczych aspektów, ale nie całości.
Np. można widzieć poszczególne kształty, ale nie rozumieć znaczenia całości.
Widzenie całości to złożony problem, wymagający koordynacji działania kory wzrokowej, ruchów oczu, skupiania uwagi, kojarzenia informacji, spójności elementów wrażeń.
Uszkodzenie pierwotnej kory wzrokowej (np. niedokrwienie) może prowadzić do czarnej dziura w polu widzenia
(mroczek, skotoma), a w rozległej formie jest to
ślepota korowa.
Ubytki w polu widzenia są często dopełniane interpolowanymi danymi, badani są przekonani, że widzą całość (podobnie jak nie widzimy obszaru plamki ślepej).
|
Uszkodzenie kory lub promienistości wzrokowej prowadzi do utraty wzroku, chociaż informacje z siatkówki trafiają do różnych części mózgu.
Ślepowidzenie (blindsight) lub ślepowzrok to szczątkowe widzenie bez wrażeń wzrokowych, które może się pojawić przy takich uszkodzeniach. Zaobserwowano je początkowo u małp, a później u ludzi (przykład: film na którym niewidomy obchodzący przeszkody). Zachowana jest częściowa zdolność do lokalizacji miejsca, ruchu, kształtu a nawet koloru, chociaż badani "nic nie widzą", tylko zgadują. Informacja dociera przez wzgórze (LGN) i wzgórek wzrokowy do wyższych pięter układu wzrokowego i płata ciemieniowego. W miarę treningu pacjenci nabierają wprawy w "wyczuwaniu" widoku, chociaż nie przypomina to wrażeń wzrokowych. |

|
|
Wrażenia wzrokowe (świadomość widzenia) pojawiają się dzięki aktywnej eksploracji, szukania informacji i rozróżnianiu stanów układu wzrokowego na poziomie skojarzeniowym, można je uznać za wewnętrzny komentarz.
|
|
Zaprzeczanie ślepocie (zespół Antona)
Pomimo ślepoty badani mają wrażenia wzrokowe, odmawiają nauki z niewidomymi, usiłują sami chodzić chociaż ciągle obijają się o przedmioty i przewracają.
Mają tendencje do konfabulacji, np. opisu widzianej osoby.
Lekceważą niezgodności opisu tworzac racjonalizacje, np. "jest noc, słabe światło, zagracony pokój".
Ślepota histeryczna to brak wrażeń wzrokowych, pomimo działającej kory wzrokowej, .
Wykrywana za pomocą odruchu obronnego, odruchu okoruchowego, badań EEG.
Zespół Charlesa Bonneta!
Proste halucynacje geometryczne pobudzają głównie V1, a twarze zakręt wrzecionowaty (FFG).
Zdeformowane twarze szczególnie pobudzają fragment FFG związany z obrazem oczu i ust (zębów); są też halucynacje związane z kreskówkami.
Halucynacje w tym zespole są raczej na niskim poziomie, wydają się jak film, zdarzają sie tylko ludziom niedowidzącym lub niewidomym, nie jest to podobne do snów bo te halucynacje nie mają jakiegoś związku z przeżyciami danej osoby.
Zespół Charlesa Bonneta opisany został w książkach Ramachandrana (Phantoms in the Brain) i Chandry (Sacred Games).
Ogólnie o halucynacjach nie tylko wzrokowych poczytać można w Wiki.

O widzeniu można się wiele dowiedzieć badając złudzenia wzrokowe.
Widzimy to, na co jesteśmy przygotowani:
Drive carefully.

Rola kontekstu w formowaniu się obrazu jest widoczna
na obrazkach pokazanych tutaj .
Lokalny pokaz wybranych złudzeń: warto zajrzeć na stronę
Akiyoshi Kitaoka.
Uwaga! Złudzenia ruchu Kitaoki mogą wywołać dezorientację i odruchy wymiotne, takie złudzenia
mają nawet koty!
Złudzenia dynamiczne: gięcie wskaźnika.
Co się stanie, jeśli ubierzemy okulary dające żółty obraz w prawym polu widzenia, a niebieski w lewym?
Zapytajcie Aline Bompas.
Sensomotoryczna teoria widzenia mówi, że ruchy oczu są niezbędnym warunkiem by nauczyć się widzieć.
Siatkówka nie ma równomiernego rozłożenia czopków, światło różnie się odbija od powierzchni, a jednak mamy wrażenia stałości kolorów.
Nastąpi dość szybka adaptacja do takiego widzenia; po 40 min. zdjęcie takich okularów powoduje, że w czasie ruchu oczu w prawo biała powierzchnia widziana jest jako niebieskawa (kompensując żółty kolor), a w lewo jako żółtawa (kompensując kolor niebieski).
Czy niewidomy może malować? Zmysł dotyku daje podobne informacje, pozwalając tworzyć wyobrażenie relacji przestrzennych, dlatego używany jest jako substytut wzroku u osób niewidomych.
Esref Armagan z Turcji, niewidomy od urodzenia, maluje używając kolorów i stosując perspektywę, np. malując oktagonalny budynek baptyserium we Florencji.
Jego kora wzrokowa reaguje na relacje przestrzenne badane przez dotyk, używane kolory często są przypadkowe, a w innych przypadkach są to wyuczone kolory znanych obiektów, np. drzewa zielone, artysta wie gdzie farby o różnych kolorach są na jego palecie (Amir Ahmed i inn, 2008).
Wyobraźnia wzrokowa związana jest prawdopodobnie z możliwościami odtworzenia w korze wzrokowej podobnych pobudzeń jak w czasie aktualnego doświadczenia.
Wymaga to dostatecznie silnego pobudzenia niższych obszarów przez wyższe obszary układu wzrokowego (podobnie jest w innych układach zmysłowych).
Dla skomplikowanych obiektów potrzebna jest przestrzeń neuronalna, w której tworzy się plan działania; bierze w tym udział kora ciemieniowa.
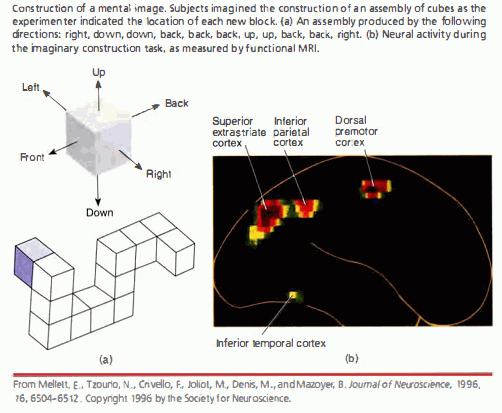
Istnieją silne indywidualne różnice w zdolności do wyobrażenia sobie i np. narysowania czegoś z pamięci (czy zaśpiewania lub zagrania melodii z pamięci).
Te różnice mierzy test żywości wyobraźni wzrokowej.
Na czym więc polega widzenie? Mechanizm ten z grubsza wygląda tak:
Układu równowagi wraz z układem czuciowym, wzrokiem i priopriocepcją bierze udział w tworzeniu mapy przestrzeni wokół ciała (peripersonalnej).
Przestrzeń bliska, w której możemy działać, jest inaczej reprezentowana niż przestrzeń odległa.
Zaburzenia reprezentacji przestrzeni peripersonalnej wywołują ciekawe efekty:
Obraz świata i relacji przestrzennych jest kombinacją egocentrycznego i allocentrycznego punktu widzenia.
Allocentryczne (od greckiego słowa "inny") postrzeganie, związane z obiektami, rozwija się pomiędzy 3 a 5 rokiem życia, kiedy dziecko zaczyna sobie wyobrażać świat z innej perspektywy niż tylko egocentryczna.
Allocentryczne postrzeganie jest skupione na obiektach, ich relacjach, własnościach, ich formie niezależnie od naszego ruchu i położenia, jest w znacznej mierze niezależne od położenia obserwatora - rozpoznajemy obiekty i znamy ich własności, chociaż im się szczegółowo nie przyglądamy, wiążemy je z pewną kategorią.
Mechanizmy przetwarzania informacji ego i allocentrycznej są częściowo znane dzięki pracom zrobiony w większości już w 21 wieku.
Allocentryczne postrzeganie wiąże się z brzusznym szlakiem wzrokowym, rozpoznawaniem obiektów jako takich, bez odwołania do swojego położenia.
Egocentryczne postrzeganie wiąże się z aktywnością prawego górnego płacika ciemieniowego (SPL), analizującego informację o tym, gdzie są w danej chwili nasze własne kończyny i jakie są ich możliwości działania uwzględniając wielkość postrzeganego obiektu.
Okolice tylnej bruzdy ciemieniowej są połączone z płatem czołowym i pełnia rolę w kontroli sakad, uwadze wzrokowej, działaniach wolicjonalnych związanych z wskazywaniem i chwytaniem.
Grzbietowy szlak wzrokowy odpowiada więc na pytanie "gdzie to jest w relacji do mnie i co mogę z tym zrobić", a nie tylko na pytanie ogólne "gdzie". Jest to konieczne by określić, czy mogę dany obiekt złapać ręką lub dosięgnąć nogą, czy warto próbować, jak ocenić głębię, siłę i miejsce uderzenia?
Uszkodzenia obszarów związanych z egocentrycznym postrzeganiem prowadzi do
zespołu nieuwagi stronnej: zanika możliwość postrzegania lewej części przestrzeni
Uszkodzenia w obrębie brzusznego szlaku - zakrętu wrzecinowatego i górnego zakrętu skroniowego w prawej półkuli - powodują zaburzenia allocentrycznego punktu widzenia - część obiektów różniących się fragmentami po jednej ze strony jest ignorowana
(Ota i inn 2001).
Widzenie wymaga więc powstania sprzężenia pomiędzy najniższymi (V1) i najwyższymi (IT, V5) obszarami układu wzrokowego.
Można to bezpośrednio stwierdzić hamując aktywność STS za pomocą
rTMS, krótkich impulsów pola magnetycznego
(Alvaro Pascual-Leone, Silvanto, Batteli i inn.); trzeba wiedzieć w którym momencie zastosować impuls (monitorować sygnały EEG).
Podsumowując: wiesz, co widzisz, i widzisz, co wiesz ...
Testy psychologiczne w latach 1950 donosiły, że ludzie mają w większości czarno-białe sny, ale w późniejszym okresie sny stały się w znacznej mierze kolorowe.
Czarno-białe sny mogły być wynikiem oglądania czarno-białej telewizji i filmów (Schwitzgebel 2002; Murzyn, 2008).
Zmiany mogą wynikać z rozpowszechnienia się kolorowej telewizji, lub szerszego używania środków farmaceutycznych.
Jednakże dane są kontrowersyjne i wygląda to tak, że wiele osób nie potrafi określić czy ma wrażenia koloru, czy nie, zapewne aktywacje ich mózgów nie przypominają dostatecznie aktywacji na jawie by można je było skategoryzować używając tych samych pojęć.
Co się stanie, gdy aktywacji ulega sam obszar IT, związany z rozpoznawaniem obiektu, a pozostałe pola wzrokowe nie są silnie pobudzone? Powinniśmy mieć wrażenie rozpoznania obiektu pomimo braku wyraźnych wrażeń kształtu czy koloru. Dowolne kombinacje są możliwe i mogą prowadzić do dziwnych wrażeń, np rozpoznania i poczucia ruchu bez koloru i kształtu.
Jeśli faza REM snu służy konsolidacji pamięci to tylko te aspekty, które są istotne powinny ulegać pobudzeniu by nie tracić niepotrzebnie energii.
Jak odróżniamy halucynacje od rzeczywistości? Nie każdy je dobrze odróżnia.
Wyglada na to, że monitorowanie co jest rzecyzwistościa zależy od aktywnosci i ukształtowania bruzdy czołowej okołoobręczy (paracingulate sulcus, PCS), w obszarze przyśrodkowo-przendiej kory przedczołowej (medial anterior prefrontal cortex, maPFC).
M. Buda et al, A Specific Brain Structural Basis for Individual Differences in Reality Monitoring, J Neurosci 2011
The findings provide evidence that individual differences in introspective abilities like reality monitoring may be associated with specific structural variability in the PFC.
Pozostałe zmysły również potrzebują kory mózgu do precyzyjnej dyskryminacji wrażeń.

|
Pierwotna kora słuchowa zajmuje pola kory skroniowej 41/42 Brodmanna, zakręt skroniowy przedni poprzeczny (zakręt Heschla), schowany w bruździe bocznej, pod stykiem płata czołowego i ciemieniowego, na płacie skroniowym.
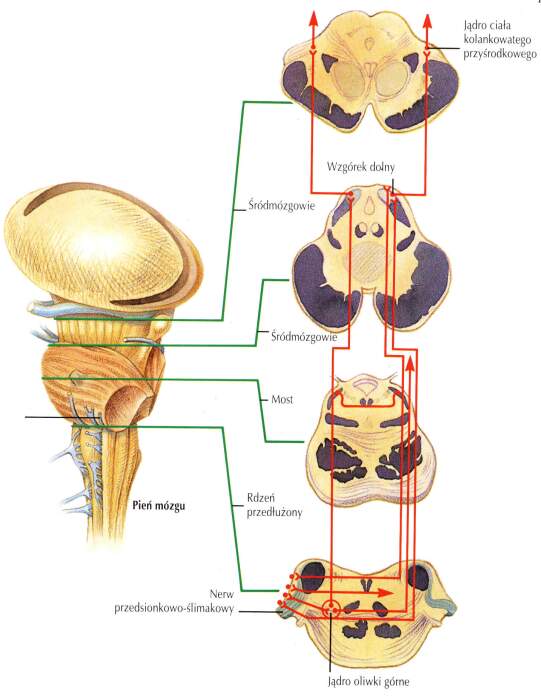
Wyspa i równina skroniowa zaangażowana jest w dalszą analizę informacji słuchowej (film, tylko lokalnie). Drogi słuchowe: ślimak ucha wewnętrznego => jądra ślimaka => jądra oliwek => ciało kolankowate przyśrodkowe (MGN, wzgórze) => promienistość słuchowa => pierwotna kora słuchowa. Drogi słuchowe (film, tylko lokalnie). Kora słuchowa kontroluje pobudliwość
przyśrodkowego ciała kolankowatego (MGN) - można naprawdę "wytężyć słuch", oczekując dźwięku z określonego kierunku lub przestać go zauważać.
Regulują to wzgórki czworacze dolne w śródmózgowiu.
|

|

|
|
|
Pobudzenia kory słuchowej dają wrażenia buczenia, stukania lub brzęczenia
(tinnitus).
Uszkodzenia kory powodują osłabienie słuchu w przeciwległym uchu, trudności z lokalizacją dźwięku.
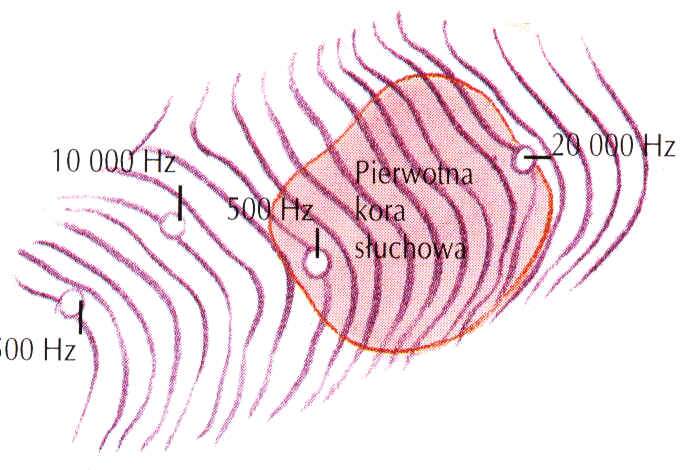
Pierwotna kora słuchowa ma budowę kolumnową, reakcje na te same częstości przy progowych dźwiękach.
Projekcja tonotopiczna - niskie częstości w bocznych, wysokie w środkowej części kory.
Złożone reakcje na podstawowe dźwięki mowy - fonemy.
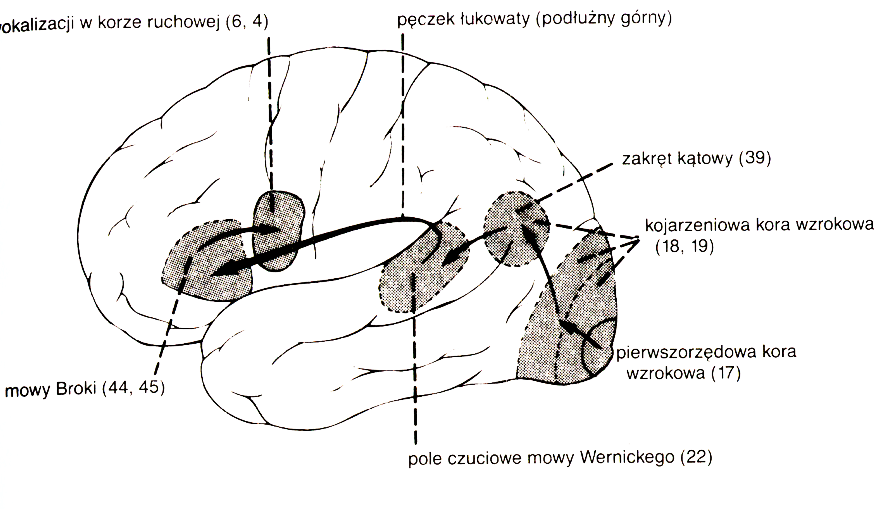
Połączenia międzykorowe:
Całkowite uszkodzenie pierwotnej kory słuchowej prowadzi do głuchoty korowej.
Wtórna kora słuchowa:
obszar Wernickego w płacie skroniowym półkuli dominującej.
Lezje wtórnej kory słuchowej wywołują różnorodne
afazje (gr. a+phasis, brak mowy).
Wczesna specjalizacja kory przy rozróżnianiu fonemów prowadzi do trudności w uczeniu się języków obcych, np. języków tonalnych.
Polakom trudności sprawia rozróżnienie np. angielskiego "thieve" i "sieve", japończykom jednakowe wydaje się "ram" i "lam".
Zdolność do rozróżniania fonemów języków obcych zanika ok. 10 miesiąca życia; jest to przyczyną trudności uczenia się języków obcych w starszym wieku: jeśli się wyraźnie nie słyszy to nie można też nauczyć się rozumieć i mówić.
Agnozja słuchowa prowadzi do upośledzenia mowy (brak rozumienia potrzeby korekcji, skoro się nie słyszy różnicy).
Rozwój map tonotopicznych jest jedną z podstaw dobrego słuchu.
Hałas, biały szum, powoduje brak zróżnicowania map w korze słuchowej u szczurów i zapewne u ludzi; sprzyja to padaczce.
Niemowlęta dorastające w hałasie mogą mieć trudności językowe, a być może nawet zaburzenia podobne do autyzmu: ich mózgi muszą się nauczyć filtrować informację i dłużej skupiać wyłącznie na jednym bodźcu, co może spowodować trudności z przenoszeniem uwagi.
Kontrastowe, proste bodźce w ciszy najlepiej rozwiną mapy słuchowe.
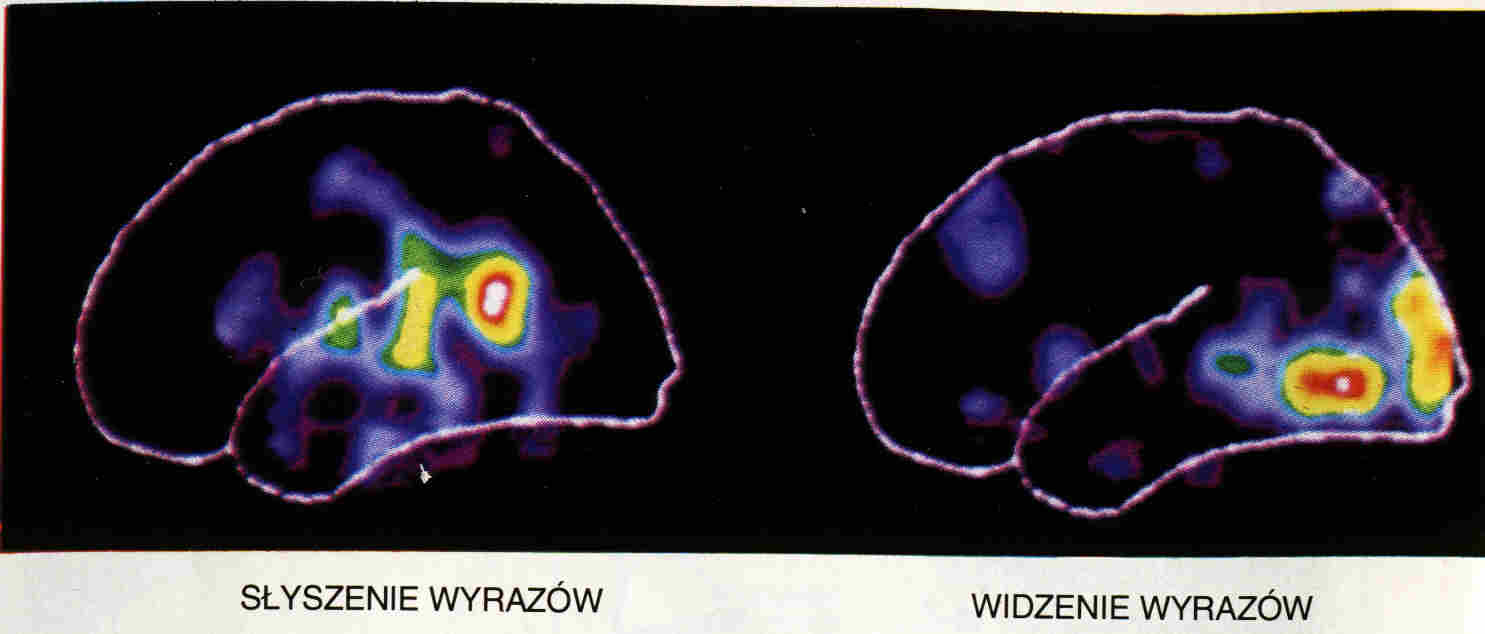
Afazja Broca (eferentna afazja ruchowa) jest wynikiem uszkodzenia obszaru Broca i prowadzi do trudności z wymówieniem słów, ciągów sylab, czasami pozostaje tylko kilka słów, które da się wymówić.
Afazja Wernickiego (akustyczno-gnostyczna afazja skroniowa), po uszkodzeniu obszaru Wernickiego, prowadzi do zaburzeń rozumienia mowy własnej i innych.
Mowa jest płynna, ale niegramatyczna, bez sensu, słowa są pomieszane, tracą znaczenie.
Niezdolność rozumieniu mowy prowadzi do niezdolności do pisania i czytania.
Pacjenci są nieświadomi swoich problemów językowych! To ładny przykład problemu, z którym zmierzyć się musi teoria świadomości.
Afazja przewodnictwa wynika z uszkodzenia pęczka łukowatego, łączącego ośrodek Wernickiego i Broca.
Mowa pozostaje płynna i rozumienie normalne, ale są trudności w powtórzeniu słowa i częste przekręcanie wyrazów, pomimo chęci skorygowania błędów.

Lezje zakrętu skroniowego środkowego dają zaburzenia pamięci słuchowo-werbalnej
(afazja akustyczno-mnestyczna).
Efekt: niezdolność do powtórzenia serii wyrazów.
Lezje tylnej części dominującego płata skroniowego:
afazje nazewnicze.
Powodem jest brak skojarzeń między wzrokowym rozpoznaniem a nazwą.
Nie można narysować przedmiotu po usłyszeniu instrukcji słownej, chociaż nie ma trudności w kopiowaniu rysunków.
Zagadnienia związane z mową związane są też z wyższymi czynnościami psychicznymi i zostaną omówione później.
Afazja akustyczna: zaburzenie rozpoznawania dźwięków niewerbalnych, odgłosów zwierząt lub sygnałów akustycznych (używana jest też nazwa
"agnozja akustyczna").
Testy: wskazać obrazek kojarzący się ze źródłem dźwięku.
Dwa rodzaje afazji akustycznej: niezdolność do odróżniania dźwięków (kora pierwotna) i niezdolność do ich kojarzenia (kora wtórna).

Interpretacja emocjonalna wrażeń słuchowych jest silnie uwarunkowana kulturowo, np. siorbanie i siąkanie japończyków i chińczyków.
Możliwe jest "słuchanie" na podstawie obserwacji ruchów warg;
Uniwersytet Gallaudeta w USA istnieje od 1857, naucza korzystając z
Amerykańskiego Języka Migowego (ASL) osoby głuche (Sacks, 1998).
Podobnie jak iluzje wzrokowe, istnieją też
iluzje słuchowe, związane z mową, dźwiękami muzyki, słuchem przestrzennym, oddziaływaniem między wzrokiem a mową.
Jest to obszerny i ciekawy temat, sporo przykładów jest na stronie
Diane Deutsch, która napisała o iluzjach akustycznych kilka książek.
Efekt Mozarta, znany z popularnej literatury, to wpływ słuchania muzyki klasycznej (od Vivaldiego i Bacha do Mozarta) na wyników testów.
Wiarygodnie pokazano pozytywny wpływ muzyki na pamięć roboczą i rozumowanie czasoprzestrzenne; nie oznacza to jednak trwałego zwiększenia inteligencji, a raczej chwilowe pobudzenie kory i polepszenie nastroju.
Nawet biały szum w słuchawkach wpływa na chwilowo lepsze wyniki w zadaniach arytmetycznych.
Nie chodzi o Mozarta tylko o muzykę pobudzającą i podnoszącą nastrój, efekt jest dość słaby, ale w pewnych warunkach zauważalny; prasa i firmy komercyjne bardzo go wyolbrzymiły.
W stanach Tennessee i Georgia każdy nowo narodzony obywatel dostaje CD z muzyką Mozarta - krytycy
kwestionują jednak, czy jest to program sensowny.
Efekty są kontrowersyjne, różnią się u ludzi starszych, dorosłych i u dzieci, zależą też od rodzaju muzyki.
Mozarta oczywiście warto słuchać, a jeszcze bardziej grać samemu, wtedy efekty będą wyraźniejsze.
Muzykoterapia ma liczne zastosowania, muzyka relaksacyjna może np. spowodować obniżenie ciśnienia i częstości napadów padaczki, stosowana jest w rehabilitacji poudarowej, stanów nerwicowych ("muzyka koi nerwy") i innych problemach.
Meloterapia to muzykoterapia śpiewem.
Choreoterapia to terapia tańcem.
Biologicznymi wpływami muzyki zajmuje się biomuzykologia; można w niej wyróżnić:

Jaka była wartość ewolucyjna muzyki?
Dlaczego słuchanie muzyki sprawia nam przyjemność?
Mowa konieczna jest do komunikacji. Do czego potrzebna była muzyka?
Steven Pinker: "Jeśli chodzi o biologiczne przyczyny i efekty muzyka jest bezużyteczna".
Czyżby muzyka nie miała ewolucyjnego sensu?
Śpiew samców ptaków, wielorybów, czy zawodzenia gibbonów służą przywabianiu samic, zwracaniu uwagi. już w starożytnych Chinach pisano:
Smutne wołania gibonów w trzech kanionach Pa-Tu,
Po trzecim ich koncercie tej nocy,
mokre od łez jest ubranie wędrowca.
Poemat Chiński z 4 wieku.
B. Merker (1999), biomuzykolog: wokalizacja małp stadnych służy komunikacji i ostrzeganiu obcych małp, jest tańsza niż ślady zapachowe.
Ksenofobia prowadzi do dziedzicznych chorób; potrzebne jest mieszanie genów.
Potrzebny jest sygnał pozwalający przekroczyć barierę strachu i przejść do innej grupy.
U szympansów (i ludów myśliwsko-zbierackich) samice wabione są głosem chóru samców.
Istniała ewolucyjna presja do wspólnej wokalizy.
Na poziomie psychologicznym związek śpiewu z szukaniem partnera, miłością i seksem jest wyraźny.
Na poziomie neurofizjologicznym śpiew pobudza ośrodki przyjemności, wyzwala mechanizm nagrody za przezwyciężenie barier między osobnikami.
Na poziomie społecznym współpraca samców zwiększała spójność wewnątrz grupy.
Piszczałki i bębny są słyszalne na większe odległości, można je używać do sygnalizacji
("mówiące" bębny tama w Afryce), były więc powody do używania instrumentów.
Pozostałe zmysły również potrzebują kory mózgu do precyzyjnej dyskryminacji wrażeń.
|
4 podstawowe smaki: słodki, słony, kwaśny, gorzki, odbierane przez różne części języka?
Kubki smakowe: około 2000-5000, każdy skupia ok. 150 receptorów, kubki są na języku i trochę na podniebieniu.
Węch jest 10.000 razy bardziej wrażliwy na stężenie cząsteczek chemicznych niż smak! Wywęszyć można pojedyncze cząsteczki.
|

|
|
Adaptacja do smaku: receptory przestają reagować, pomimo ciągłego pobudzania.
Kora smakowa mieści się w zagłębieniu płata ciemieniowego, okolicach zakrętu zaśrodkowego, niedaleko kory somatosensorycznej SI reprezentującej język.
Zespół smakosza (Gourmand syndrome): uszkodzenie prawego płata czołowego, związanego z układem nagrody, może wywołać obsesję na punkcie wykwintnego jedzenia.
|

|
Ludzie mają około 40 mln komórek węchowych w obszarze około 2 na 5 cm; psy około 1 mld, a najlepszy węch mają niedźwiedzie (oceniany na 7 razy lepszy niż psy).
Znamy około 1000 różnych
receptorów węchowych,
ale każdy zapach pobudza wiele z nich i każdy receptor reaguje na wiele zapachów.
Kodowanie zapachów jest kombinatoryczne, podobnie jak słowa zapisywane za pomocą liter różne wrażenia zapachowe związane są z pobudzaniem kombinacji receptorów (mechanizm ten częściowo poznano w 1999 roku).
U człowieka ponad 400 genów koduje białka receptorów węchowych.
Kobiety mają średnio lepszy węch, ale są duże indywidualne różnice.
Ok. 5 mln neuronów przesyła sygnały przez nerw węchowy do skupisk neuronów w opuszce węchowej, zewnętrznym, pierwotnym fragmencie kory (neurony opuszki podobne są do komórek nabłonka skórnego).
Węch doprowadza informacje bezpośrednio do ciała migdałowatego układu limbicznego => szybkie reakcje emocjonalne, strach wywołany zapachem.
Uszkodzenia kory węchowej powodują nieprzyjemne halucynacje węchowe, czasami to oznaki ataku padaczki.
Drogi pobudzeń (Kolb & Whishaw, 1996):

Wtórna kora węchowa:
hak (brzuszna część kory czołowej) i
kora śródwęchowa, bliska formacji hipokampa.
Jedyny system, który nie łączy się z korą wyłącznie przez wzgórze; większość kory węchowej to kora stara, 3-warstwowa, ale kora śródwęchowa ma 6 warstw.
Interpretacja wrażeń węchowych jest bardzo indywidualna, zależna od wspomnień, często niejasna - pobudzenie struktur limbicznych i niespecyficzne pobudzenia różnych okolic kory.
Kora oczodołowa jest obszarem, w którym schodzą się informacje o smaku, zapachu, dotyku i rozpoznaniu wzrokowym, ma silne projekcje do prążkowia i bocznej części podwzgórza.
Ageuzja, hypogeuzja, dysgeuzja, i hypergeuzja to brak, redukcja, zniekształcenie i zwiększona wrażliwość smakowa.
Wrodzona anosmia. skorelowana jest z wrodzoną analgezją (brakiem odczuwania bólu), dochodzi do niej z powodu mutacji genu SCN9A, którego białka tworzą sodowy kanał jonowy Nav1.7 w receptorach bólowych (nocyceptorach) jak i receptorach węchowych.

Informacje na temat sygnałów z narządów zmysłów dochodzą przez 12 par
nerwów czaszkowych,
informacje węchowe przez nerw I.
Nerw zerowy (terminalny) leży bardzo blisko nerwu I, odkryty w 1913 ale zwykle nie wymieniany w podręcznikach anatomii.
Prawdopodobnie podłączony jest do ludzkiego odpowiednika
narządu Jakobsona (narządu przylemieszowego), który u wielu zwierząt ma receptory
feromonów i dopiero w latach 1990 został jednoznacznie opisany u ludzi.
Funkcje narządu przylemieszowego są nadal kontrowersyjne, nie łączy się on z opuszką węchową.
Grupa komórek w jamie nosowej łączy się z
jądrami przegrody oraz
jądrem przedwzrokowym podwzgórza (preoptic area), które powiązane są z reakcjami seksualnymi.
Wiadomo, że geny odpowiedzialne za tworzenie receptorów feromonów u myszy są również u człowieka (Linda Buck, Stephen Liberles, 2006).
Niewielka część połączeń nerwu trafia do siatkówki: długość dnia jest ważnym parametrem sterującym zachowaniami seksualnymi.
Przerwanie nerwu zerowego u chomików powoduje utratę zainteresowania partnerem.
Elektryczna stymulacja u różnych zwierząt wywołuje reakcje seksualne, np. uwalnianie spermy u złotych rybek.
U płaszczek zakończenie nerwu zerowego w podwzgórzu uwalnia hormony regulujące zachowania seksualne (np.
GnRH, R.D. Fields, Univ. Maryland).
Wieloryby i delfiny nie mają węchu (nos zastąpiła im dziura oddechowa na grzbiecie), ale mają nerw zerowy.
Kobietom podobają się bardziej zapachy koszul noszonych przez mężczyzn o odmiennym układzie immunologicznym, ale zażywanie pigułek antykoncepcyjnych zmienia te preferencje.
Środki antykoncepcyjne i zapachowe mogą zaburzać normalne działanie systemu doboru partnera, w efekcie mogą się przyczyniać do wzrostu liczby niedobranych par.
Aromaterapia, czyli leczenie zapachami, oparte jest na sensownym założeniu, że zapachy mogą wywoływać reakcje fizjologiczne.
Nie ma jednak dowodów na specyficzne działanie terapeutyczne; zapachy mogą nam zmieniać nastrój, ale działania uzdrawiajace to wynik efektu placebo.
Łzy wydają się bezwonne, jednak pokazano obniżenie się poziomu pobudzenia seksualnego (reakcje fizjologiczne, poziom testosteronu) u mężczyzn w reakcji na łzy kobiet (Shani Gelstein, Noam Sobel, 2011). Brak jest jeszcze badań wpływu męskich łez na kobiety (mężczyźni rzadziej płaczą, więc badania są trudniejsze).
Łzy prawdopodobnie zawierają feromony.

Zachowanie równowagi wymaga współdziałania
Skomplikowany system, wiele struktur.
Odruchy równowagi powstają dzięki móżdżkowi; kot zwykle spada na cztery łapy.
Odruch przedsionkowo-oczny (VOR) jest konieczny do prawidłowego widzenia.
|
Tylna część kory wyspy i część
płata ciemieniowego otrzymuje sygnały z układu zachowania równowagi.
Wzrok może dostarczać wystarczającej informacji dla orientacji.
|
 
|
Układu równowagi wraz z układem czuciowym, wzrokiem i priopriocepcją bierze udział w tworzeniu mapy przestrzeni wokół ciała (peripersonalnej).
Przestrzeń bliska, w której możemy działać, jest inaczej reprezentowana niż przestrzeń odległa.
Zaburzenia reprezentacji przestrzeni peripersonalnej wywołują ciekawe efekty:

Stymulacje elektryczne prawej półkuli w okolicach zakrętu kątowego mogą wywołać różne wrażenia autoskopowe:

Inne zaburzenia relacji przestrzennych:
Autotopagnozja wiąże się z lezjami lewego płata ciemieniowego, pacjenci nie mają problemu z wskazywaniem i dotykaniem np. części maszyn;
nie mogą też narysować czy prawidłowo poskładać układankę przedstawiającą części ciała (inna nazwa to somatotopagnozja, Gerstmann 1942), podkreślająca, że jest to niezdolność do prawidłowego postrzegania ciała.
Autotopagnozja często występuje razem z heterotopagnozją, ale znane są nieliczne przypadki czystej heterotopagnozji (lezje górnej kory ciemieniowej) i czystej autopagnozji (lezje dolnej kory ciemieniowej).
Pacjenci z heterotopagnozją nie potrafią wskazać (funkcja komunikacyjna, wskazywanie na kogoś), a potrafią uchwycić (działanie bez komunikacji).
mogą też w niektórych przypadkach wskazywać części ciała innych osób zamiast swoich (Langavant i inn, 2009) oraz dotknąć odpowiedniej części, a nie wskazać. Są to rzadkie przypadki i nie ma tu jednoznacznego opisu.
Są też liczne inne zaburzenia związane z zaburzeniem postrzegania ciała, np. autoprosopagnozja, niezdolność do rozpoznawania własnej twarzy (Vignemont 2009).
Utrzymanie homeostazy wymaga postrzegania stanu wewnętrznego organizmu, uzupełniania zasobów (tlenu, płynów, pożywienia, regeneracji).
Poczucie pragnienia i głodu można uznać za zmysł wewnętrzny.
Pragnienie: organizm składa się w 70-75% z wody i jej poziom utrzymywany jest przez mechanizmy homeostazy z dokładnością do 0.25%.
Poczucie pragnienia wymaga pomiaru: robią to
osmoreceptory w podwzgórzu
(jądro OVLT) oraz
narządzie podsklepieniowym.
Komórki te reagują na
ciśninie osmotyczne, czyli różnicę ciśnień wynikających z różnic stężeń substancji chemicznych wewnątrz i na zewnątrz błony komórek.
Pobudzenia tych komórek są wynikiem utraty wody (na skutek wydalania, pocenia się, oddychania), głównie zmian lepkości i składu
osocza krwi.
Pojawia się suchość w ustach, gdyż
gruczoły ślinowe otrzymują wodę z krwi.
Jednocześnie tylny płat przysadki mózgowej wydziela
hormon antydiuretyczny (wazopresyna),
powodując resorpcję wody i jonów sodu w kanalikach nerkowych.
Zmiany własności osocza są powolne, dlatego istnieje szybszy mechanizm informowania mózgu i wygaszania pragnienia: reakcja na zwilżanie i ochładzanie jamy gębowej i gardła; dlatego pijemy więcej wody ciepłej niż zimnej. Zwilżanie obniża pragnienie na 30-40 minut, w tym czasie powinny zadziałać powolne mechanizmy informujące o wzroście poziomu płynów.
Są również dodatkowe receptory w żołądku.
Podobnie jak mamy zaburzenia percepcji, tak i są różne zaburzenia poczucia pragnienia.
Głód: masa ciała jest zwykle bardzo stabilna.
Karl Lashley (1938) podejrzewał, że głód nie jest reakcją na pusty żołądek.
Wycinając różne fragmenty mózgu szczurom pokazał, że główną rolę w poczuciu głodu gra podwzgórze.
Ośrodek oceniający poziom głodu można zlokalizować w jądrach bocznych podwzgórza, a ośrodek oceny sytości w jądrze brzuszno-przyśrodkowym.
Podwzgórze reaguje na poziom hormonów takich jak
leptyna, wydzielana przez komórki tłuszczowe,
grelina wydzielana w żołądku, przewodzie pokarmowym oraz
jądrze łukowatym podwzgórza (ARC), które integruje sygnały regulujące łaknienie wydzielając neuropeptydy
NPY
i
AGRP.
Leptyna wydzielana jest do krwi przez komórki tłuszczowe, informując podwzgórze, że organizm ma już wystarczająco dużo kalorii; kontrola poziomu leptyny pomaga w regulacji normalnego odżywiania.
Spadek poziomu leptyny powoduje wzrost poziomu NPY i AGRP poczucie głodu, którego zadaniem jest zmusić organizm do poszukiwania pożywienia.
NPY aktywuje wydzielanie greliny, poziom zmienia się szybko, w ciągu minut; AGRP działa znacznie wolniej, wstrzyknięcie tego peptydu szczurowi powoduje obiadanie się przez parę dni.
Niedobory białka, tłuszczy, cukrów (glukozy we krwi), witamin czy soli mineralnych mają również wpływ na łaknienie i stwarzają motywację do spożywania specyficznych pokarmów.
Zaburzenia mechanizmu łaknienia
prowadzą do jadłowstrętu (anoreksji), żarłoczności psychicznej (bulimii) i innych chorób.
Łaknienie spaczone jest bardziej złożonym zaburzeniem, występuje względnie często u kobiet w ciąży, polega na konsumpcji różnych niejadalnych substancji, w tym normalnie wywołujących poczucie wstrętu.
Mechanizm jest skomplikowany i poznany tylko częściowo.
Dobra strona o energii dla mózgu oraz indeks glikemiczny, wskazujący na poziom glukozy we krwi w parę 3 godzin po spożyciu pokarmu.
Ssaki są stałocieplne. Termoregulacja wymaga termoreceptorów i mechanizmu regulacji wydzielania ciepła.
Liczne inne funkcje homeostatyczne odbywają się całkowicie poza świadomością.
Percepcja upływu czasu jest szczególnym rodzajem postrzegania. Trzeba tu odróżnić czas fizyczny (mierzony przez zegary), neuronalny (różne okienka integracji informacji) oraz psychologiczny.
Procesy zachodzące regularnie, takie jak rytmy okołodobowe, postrzegane są odmiennie od szybkich zmian.
Powtarzające się bodźce ulegają habituacji i postrzegana długość zdarzeń wydaje się krótsza, nowe bodźce wymagają więcej energii i dłuższego czasu analizy, wydają się subiektywnie dłuższe.
Zmysły nie działają w izolacji od siebie, w móżgu wszystko się ze sobą silnie łączy, postrzeganie służy działaniu.
Działamy tak, jak nam na to pozwala aktywność neuronalna, mapowanie sesno-motoryczne, percepcja pozwalająca dostrzegać okazję do działania (afordancje), chwytania, manipulacji przedmiotami.
Orientacja, postrzeganie przestrzeni i afordancji wymaga integracji zmysłów i układu ruchu.
Integracja wielomodalna: na co możemy zwrócić uwagę? Czego się dowiedzieć o stanie swojego mózgu?
Nie potrafimy odizolować wrażenia kształtu, ruchu czy koloru od całościowego postrzegania, ale możemy skupić się na jednym z aspektów wrażeń wzrokowych lub na całościowym doświadczeniu.
Czy możemy skupić się i wiedzieć co dzieje się w dowolnym obszarze mózgu?
Czy nadanie nazw specyficznym aspektom doświadczenia ułatwi nam takie skupianie?
Dobre przykłady? Wrażenia słuchowe osób niewidomych, o których pisze Sacks.

Substytucja zmysłów to ogólna technika używania zmysłów jako receptorów wrażeń o różnych modalnościach.
Substytuty orientacji przestrzennej: sonifikacja sygnałów z kamery, zamiana na wibracje odczuwalne przez skórę, zamiana na bodźce stymulujące język.
Postrzeganie pola magnetycznego za pomocą wibracji pozwala na wyrobienie sobie nowego "zmysłu magnetycznego", podobnie z polem elektrycznym - rekiny używają go do wyszukiwania ofiar.
Stymulacja języka informacją z kamery prowadzi do powstania najpierw chaotycznych i niestabilnych wrażeń.
Jak powstają nowe wrażenia i jakie to są wrażenia (qualia)?
Mózg uczy się pamiętając powtarzające się wzorce, porównuje zapamiętane z aktualnymi, dodaje kontekst, cała reakcja i możliwość interakcji prowadzi do powstania specyficznych wrażeń.
Kevin O'Regan opisuje np. wrażenie związane z miękkością gąbki, które wymaga interpretacji działania, dotyku (konieczny jest nacisk na powierzchnię), poczucia sprężystości będącego reakcją na silniejszy nacisk.
Nie chodzi tylko o to co robimy, ale co możemy zrobić i co pamiętamy, o "osiągalność" (affordance) pewnych działań i odpowiadających im stanów mózgu.
Nieliczne osoby niewidome nauczyły się posługiwać echolokacją, wywołując odbite fale kliknięciami języka lub uderzeniami laski (Thaler i inn. 2011).
U osób niewidomych aktywna była bruzda ostrogowa (calcarine sulcus) w płacie potylicznym, obszar kory wzrokowej V1.
Pod wieloma względami możliwe są całkiem nowe qualia:
Każdy odczuwa świat w nieco inny sposób.
Silne kojarzenie bodźców zmysłowych o różnych modalnościach to synestezje (Gr. syn = razem + aisthesis = postrzegać); nazwę wymyślił
Francis Galton,
Smak lub poza nazwy, widok lub smak dźwięku, czucie widoku, kolory liter ... wszystkie kombinacje.
Wladimir Nabokov - doskonały przypadek kolorowego słuchu.
Długie aaa - odcień starego drewna, g to wulkanizowana guma, k jest jagodowe ...


Zjawisko rzadkie, dla 2 modalności 1:25 000.
Jest przynajmniej 39 rodzajów synestezji!
Najczęściej: wzrok + słuch w formie leksykalno-kolorowej: litery i liczby widziane i słyszane są w kolorach, ok. 2/3 wszystkich przypadków.
Wrażenie koloru może być wywołane przez widok grafemów, cyfr zegara, dźwięków muzyki, ogólnych dźwięków, fonemów, nut, zapachów, smaku, bólu, osobowość rozmówcy, dotyk, temperaturę, podniecenie, emocje.
Bardzo rzadko w synestezjach występuje dotyk i smak, najrzadziej węch.
U kobiet zdarza się kilka razy częściej niż u mężczyzn (3-8 razy w różnych badaniach).
Przejściowe synestezje: wywołane anty-serotoninergiczny środkami halucynogennymi, deprywacją sensoryczną, stymulacją elektryczną lub padaczką skroniową, urazami głowy.

Czym jest wrażenie? Fala świetlna, wibracja mechaniczna czy pobudzenie chemiczne => podobne impulsy elektryczne.
Wrażenia wzrokowe: generyczne, konkretne kształty spiral, siatek, błyszczących ruchomych kropek, stosunkowo proste pobudzenia układu wzrokowego na poziomie V1.
Kolory dotyczą muzyki i mowy, ale wrażenie może być też zależne od ortografii.
Synestezje nie zawsze są przyjemne: usłyszenie "pięć" wywołuje wrażenie cyfry 5 na szarym tle i gwałtowne skurcze prawej części twarzy.
Obrazowanie mózgu pokazuje silne pobudzenia obszarów kory sprzężonych zmysłów.
Wzmożona aktywność struktur limbicznych, obniżona kory - jak u niemowląt.
Hipotezy:
Elizabeth Sulser z Zurichu ma wyjątkowo silne połączenia pomiędzy korą smakową, słuchową i wzrokową.
Podział informacji zmysłowej prawdopodobnie umożliwia większą precyzję działania.
Teoria rozwojowa: niemowlęta do 4 miesiąca doświadczają synestezji? Mowa pobudza wówczas potencjały wywołane nie tylko w płatach skroniowych ale i potylicznych.
Do pewnego stopnia każdy ma synestezje, np. polecenia werbalne lub dźwięki mogą w nas wywołać wyobrażenia wzrokowe.
Wielu ludzi w podobny sposób kojarzy ze sobą zapachy, kolory, wrażenia dotykowe, a nawet kolory i kształty, mamy więc do czynienia z podprogową synestezją.
Przykład: który obrazek nazwiemy
Buba a który Kiki?

Prezentacje studentów: Magdalena Kiełbasiewicz, Karolina Kmieciak, Magdalena Łaszewska, Martyna Soborska.
Schematy dla zmysłów (PPT)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}